
The Cytokine IL-12 Protects Against Neuroinflammation by Signaling to Neurons
Post by Trisha Vaidyanathan
The takeaway
The cytokine interleukin-12 (IL-12) has a protective effect in neuroinflammatory diseases. The neuroprotective effect of IL-12 is mediated by neuronal sensing of IL-12 via the IL-12 receptor, resulting in transcriptional changes that increase survival, proliferation, and protection.
What's the science?
Central nervous system autoimmune disorders like multiple sclerosis (MS) are characterized by cytokine dysregulation, in which the body’s own immune system attacks tissue, causing inflammation that ultimately damages the brain. The cytokine IL-12 is a well-established driver of this type of inflammation. Paradoxically, mice lacking IL-12 develop worse neuroinflammation and clinical outcomes in several disease models, suggesting IL-12 also has a protective role. This week in Nature Neuroscience, Andreadou, Ingelfinger, and colleagues address this paradox by investigating the hypothesis that IL-12 elicits different effects depending on the cell type receiving the signal.
How did they do it?
First, the authors asked which cells are necessary to mediate the neuroprotective effect of IL-12. Using the cre-lox system, the authors were able to delete the IL-12 receptor from specific cell populations in a mouse model of MS (the EAE model) and investigate which cell-type specific deletion altered clinical outcomes.
Second, the authors used immunohistochemistry and RNA-fluorescence in situ hybridization to visualize IL-12 receptors and IL-12 receptor mRNA transcripts, which allowed them to identify which cell types in the brain are capable of sensing IL-12. Additionally, the authors confirmed their findings from the mouse in humans by analyzing human RNA-sequencing datasets and performing immunohistochemistry in MS patient tissue.
Third, the authors performed single nuclei RNA sequencing on mice where the IL-12 receptor was deleted in neurons and oligodendrocytes in order to reveal several cell-type specific transcriptional changes induced by the absence of the IL-12 receptor. Lastly, the authors used a neuronal cell culture system to test the effect of IL-12 on isolated mouse neurons and confirm their findings on the role of neuronal IL-12 signaling in neuroprotection.
What did they find?
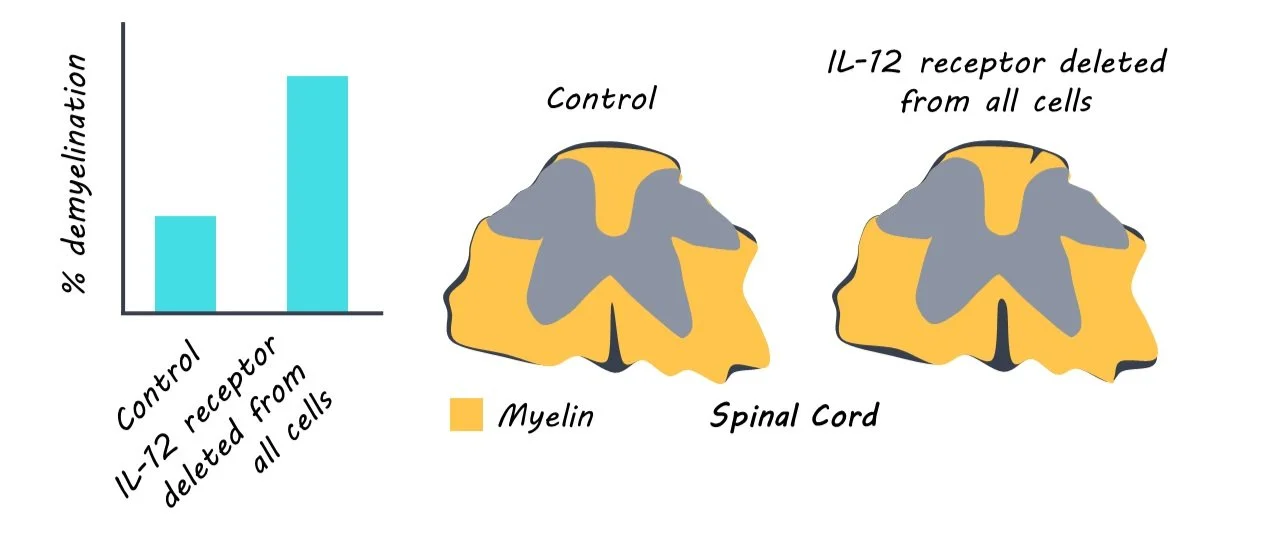
First, the authors deleted the IL-12 receptor from all cells and found the mice exhibited worse MS clinical symptoms, confirming previous findings that IL-12 can be protective. Next, the authors demonstrated that the protective effect was not driven by the sensing of IL-12 in immune cells because deleting the IL-12 receptor in all bone-marrow-derived immune cells, just T-cells, or just natural killer (NK) cells, did not change clinical disease symptoms in the mice.
Second, the authors demonstrated that both neurons and oligodendrocytes in the mouse brain express the IL-12 receptor. These findings were also confirmed in human MS patient tissue and sequencing datasets. Deleting the IL-12 receptor in neurons and oligodendrocytes led to worse disease symptoms and neuroinflammation, revealing that the neuroprotective effect was dependent on these cells.
Third, the authors answered the question of what IL-12 signaling in the brain does. The deletion of the IL-12 receptor in neurons and oligodendrocytes led to several transcriptional changes across many cell types. Overall, they found a reduction in gene expression for neuroprotection and survival, as well as a broad class of proteins called trophic factors released by cells to protect and support other cells.
Lastly, the authors narrowed in to identify that neuroprotection is mediated by neurons, not oligodendrocytes: When the authors deleted the IL-12 receptor only in oligodendrocytes there was no change in the MS clinical symptoms. To confirm this, the authors administered IL-12 to a cell culture system where only mouse neurons were present and found IL-12 increased the expression of genes that promote neuroprotection and remyelination as well as induced the release of trophic factors that are important for oligodendrocyte survival and function.
What's the impact?
This study is the first to settle the paradox of IL-12 by demonstrating that IL-12, a well-established driver of inflammation, can protect against inflammation by signaling to neurons. Understanding the protective effect of IL-12 can provide critical insight for clinical interventions in autoimmune disorders or neurodegenerative diseases that cause neuroinflammation.




