Neural Activity in Subcortical Regions is Highly Correlated with Resting State Cortical Network Dynamics
Post by Meredith McCarty
The takeaway
Despite the high interconnectedness of cortical and subcortical regions, the subcortex remains an understudied area of the human brain. High-resolution fMRI reveals that the subcortex is highly functionally connected (i.e. a high degree of correlated brain activity) with distinct cortical regions, demonstrating a gradient of connectivity for both integrative and segregated patterns of information processing.
What's the science?
In daily life, humans integrate information constantly, a process thought to be orchestrated by a highly interconnected brain. The brain is composed of many cortical and subcortical structures (i.e. neural formations deep within the brain), and there are complex patterns of connectivity between these regions that enable rapid network dynamics to unfold. To study these dynamics in healthy subjects, researchers can utilize noninvasive imaging techniques, although these methods are often limited in resolution to recording from the cortex. The lack of access to subcortical structures occurs primarily due to tooling and analysis limitations, including low signal-to-noise ratios and the varying properties of brain tissue in deeper structures. Because of these limitations, it is unknown how subcortical regions participate in neural information processing at a network level. This week in The Journal of Neuroscience, Groot and colleagues utilize high-resolution fMRI imaging techniques to measure the level of correlated activity between cortical and subcortical structures in humans.
How did they do it?
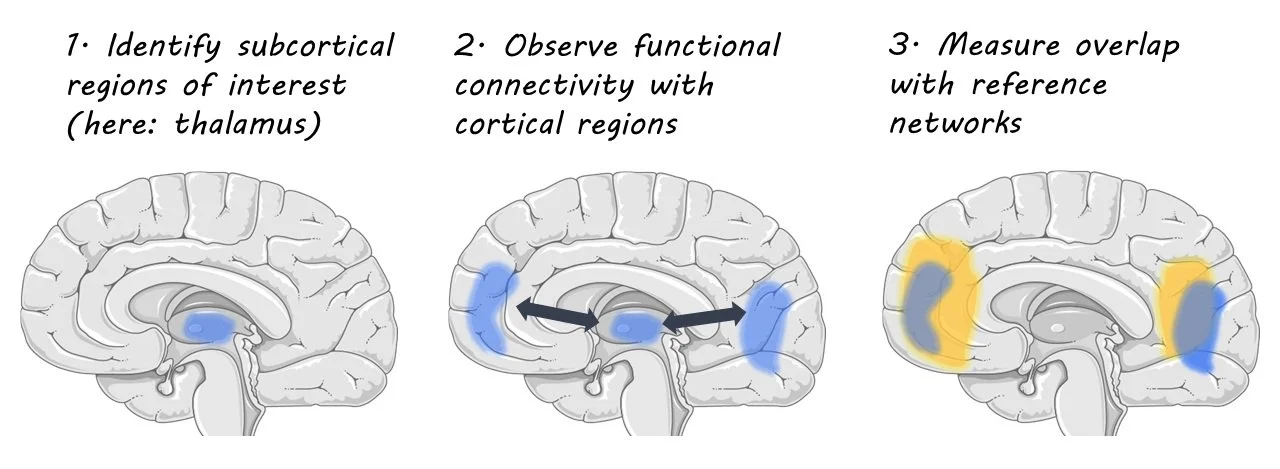
The authors utilized functional magnetic resonance imaging (fMRI) at a high-field resolution in order to record changes in BOLD signal across cortical and subcortical regions. They recruited 40 adults (21f) to participate in wakeful rest fMRI data collection, which consisted of two 15-minute sessions during which participants fixed their gaze on a central fixation point. They next determined cortical and subcortical regions of interest through use of automated parcellation algorithms. Their method of fMRI data analysis enabled the identification of patterns of intrinsic functional connectivity (FC) in the brain at a resolution that revealed more subtle functional organization. They compared subcortical FC patterns data with ‘reference networks’; common patterns of resting-state fMRI activation in the cortex known to be highly correlated with various behavioral and cognitive activities. These include the salience, visual, and default mode networks. By comparing the overlap between areas with FC to subcortical regions and the reference networks, the authors were able to measure the degree of network correlation between cortical and subcortical regions.
What did they find?
First, the authors found 7 subcortical regions that had high spatial correlation between brain regions they were functionally connected to and reference networks. These regions were the Thalamus, Striatum, Claustrum, Globus pallidus external, Hippocampus, Ventral tegmental area, and Substantia nigra. The authors dub the patterns of network correlation discovered to be “echoes” of intrinsic connectivity networks within cortical regions. Upon closer examination, they found a heterogeneous organization of echoes within subcortical subregions, with some regions exhibiting high correlation with many reference networks, while other regions exhibited very little to no significant connectivity. This suggests that there is a gradient within subcortical regions of distinct and integrated network dynamics.
What's the impact?
This study found activity in distinct subcortical regions to be highly correlated with distinct resting state cortical networks. These results suggest that the subcortex is involved in integrated multi-network neural activity with many cortical regions, an area of research previously restricted by methodological constraints. Altered subcortical dynamics are linked to many brain disorders, therefore a greater understanding of the role of subcortical regions in resting state healthy brain dynamics is pivotal.