
A Neural Prosthetic to Improve Memory
Post by Trisha Vaidyanathan
The takeaway
A neural prosthetic in the hippocampus has the potential to improve memory. In Alzheimer’s disease patients, using a personalized prosthetic to stimulate neurons in specific patterns improved memory performance in some individuals.
What's the science?
The ability to selectively preserve important or urgent memories can have major therapeutic benefits for patients who suffer from memory loss in diseases like Alzheimer’s Disease. Neural prosthetics that can stimulate the hippocampus, the brain area most associated with memory, have exciting potential. However, existing approaches have relied on stimulation to try to boost general memory function and the underlying biological mechanism is unclear. This week in Frontiers in Computational Neuroscience, Roeder, a research fellow in the department of translational neuroscience at Wake Forest University School of Medicine, and colleagues at Wake Forest and the University of Southern California tested the effectiveness of a new neural prosthetic approach to improving memory.
How did they do it?
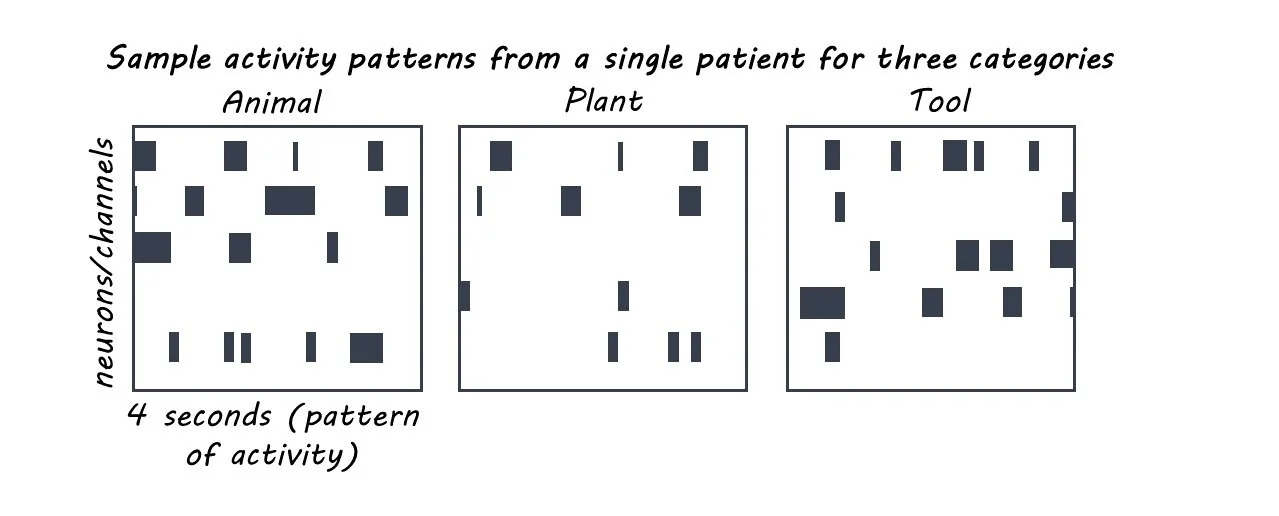
Fourteen adult subjects underwent surgery for hippocampus implants that allowed for continuous recording and stimulation of single-neuron activity. After surgery, all subjects performed a “Delayed Match to Sample Task” in which they were shown a “sample image”, followed by a 3-5 second delay, and then asked to identify the sample image amongst a choice of four images. The sample images fell under one of 5 categories: animal, building, plant, tool, or vehicle.
Next, the authors used the neural recordings obtained during the Delayed Match to Sample Task to identify the spatiotemporal pattern of neuronal activity elicited by each category of sample image. For each individual patient, a computer model was used to calculate a stimulation pattern for each image category that was derived from their neural response to all images within that category. The result was five fixed patterns of stimulation, uniquely designed for each patient, corresponding to each of the five image categories (animal, building, plant, tool, vehicle).
Finally, the subjects repeated the Delayed Match to Sample Task but during the sample image presentation, they were exposed to either (1) no stimulation, (2) “match stimulation”, in which the implanted prosthetic stimulated neurons in the pattern intended to match the category of the sample image, (3) “non-match stimulation” in which the stimulation corresponded to a category that did not match the sample image, or (4) random stimulation. After 15-20 minutes, the effect of stimulation on memory was tested by administering a Delayed Recognition Task to the subjects, in which they were asked to rank the familiarity of a collection of images that included the previous sample image. A correct trial was one in which the sample image was ranked as greater than a 2 out of 5 in familiarity, and the ranking was equal to or higher than the other image options.
What did they find?
When performance was examined for each image category for “match stimulation,” instances of increased performance occurred about twice as frequently as instances of decreased performance. Further, enhanced memory performance was primarily observed when the patient received bilateral stimulation, rather than unilateral stimulation, and had pre-existing memory impairments. This suggests that bilateral stimulation in select patients with pre-existing impairments might have the most therapeutic potential.
In trials where subjects received a “non-match stimulation,” the authors observed more instances of decreased performance relative to trials with no stimulation or match stimulation. This supported the authors’ prediction that “non-match stimulation” would interfere with natural memory processes. Overall, however, the result on memory was variable across patients and image categories and suggested that the stimulation patterns created by the computer model may not have been as specific to the image category as intended.
What's the impact?
This study provides exciting evidence to support the use of neural prosthetics in the hippocampus for enhancing memory and identifies several areas for further development. Therapeutic approaches like these may have dramatic impacts on patients who suffer from memory loss, such as those with Alzheimer’s disease.


